Title:
Take home message:
(1)In addition to membrane tension restricted to the plasma membrane, cell volume changes also have additional effects spreading all over a cell, such as changing intracellular molecular crowding states. 除了限制在质膜上的膜张力之外,细胞体积的变化还具有遍布整个细胞的额外效应,例如改变细胞内的分子拥挤状态。
(2)This ICS is land plant–specific and does not exist in yeast, algae, or animal DCP5 homologs. On the other aspect, by uncovering mRNA and translation regulators as its components, DCP5 condensate凝结物 was identified as a distinct type of plant stress granule, which we designated as DCP5-enriched osmotic stress granule (DOSG). 这种ICS是陆地植物特有的,不存在于酵母、藻类或动物DCP5同系物中。另一方面,通过发现mRNA和翻译调节因子作为其成分,DCP5冷凝物被鉴定为一种独特类型的植物胁迫颗粒,我们将其命名为富含DCP5的渗透胁迫颗粒(DOSG)。
(3)植物DCP5蛋白中特有的拥挤传感器(ICS) ,引起DCP5相分离,将翻译组和转录组从生长促进改为应激适应。
Main:
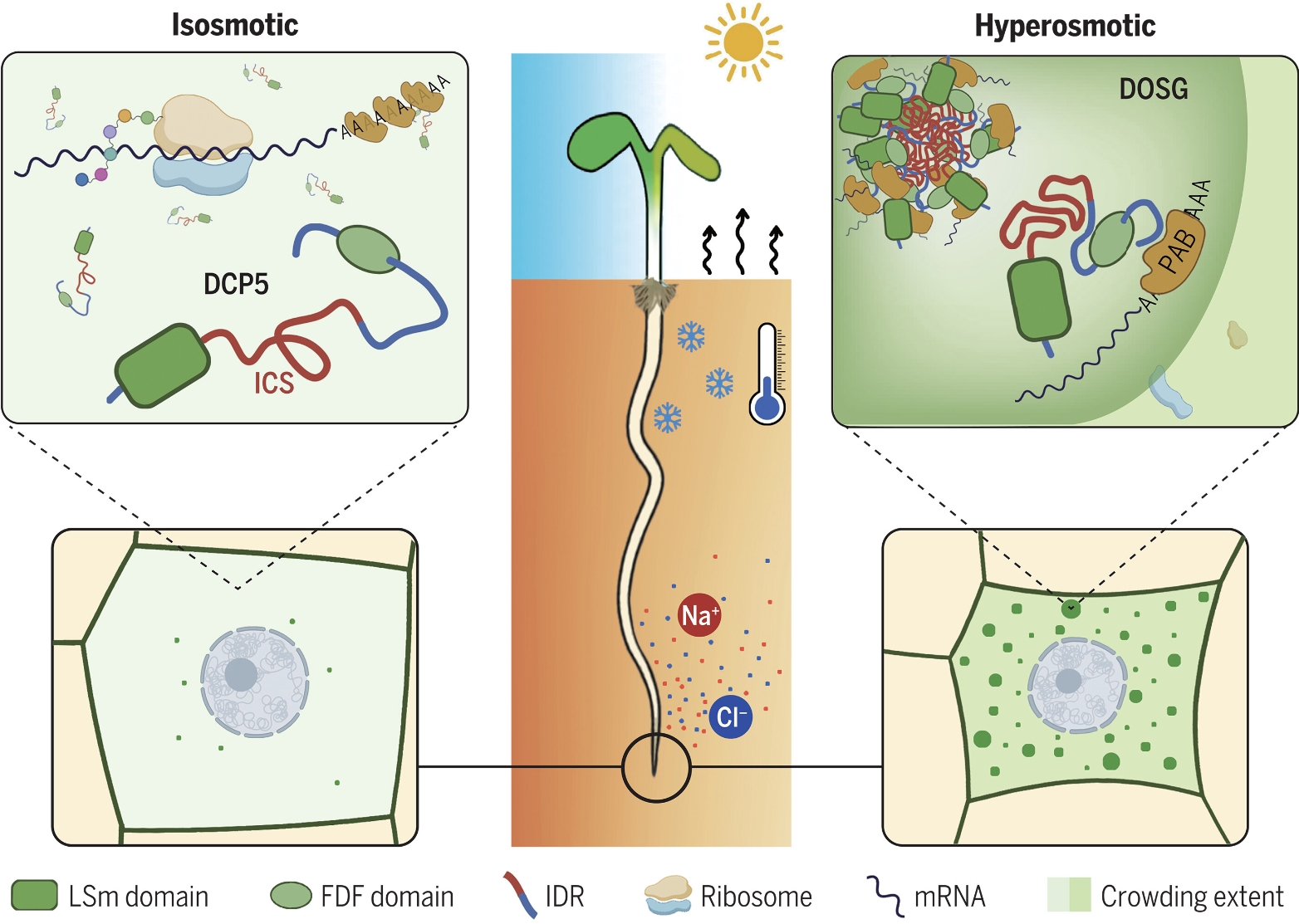
一种由DCP5介导的细胞质渗透感应机制。
(左)在等渗条件下,植物细胞在体积上是稳定的,而DCP5通常是扩散的。(右)一旦暴露于高渗状态,细胞收缩DCP5 ICS感知到分子拥挤加剧,进而触发DCP5相分离和DOSG组装。通过隔离mRNAs和调节蛋白,DOSGs适应性地重新编程翻译组和转录组,以帮助胁迫耐受性。
Editor’s summary
(1)Plants are continuously exposed to osmotic stress conditions, but how they sense osmolarity渗透压changes is not well understood. 植物持续暴露于渗透胁迫条件下,但它们如何感知渗透压的变化还不清楚。
(2)Wang et al. report that the Arabidopsis DECAPPING 5 (DCP5) protein functions as an osmosensor渗透感受器 for extracellular hyperosmolarity高渗性. Wang等人报道了拟南芥去盖蛋白5 (DCP5)作为细胞外高渗的渗透传感器。
(3)This function is achieved through molecular crowding–triggered phase separation in which the DCP5 protein undergoes a conformational change to drive phase separation. 这一功能是通过分子拥挤触发的相分离实现的,其中DCP5蛋白经历构象变化以驱动相分离。
(4)DCP5 reversibly assembles to DCP5-enriched osmotic stress granules 微粒upon hyperosmolarity and sequesters 隔离mRNA and regulatory proteins. 在高渗状态下,DCP5可逆地装配成富含DCP5的渗透应激颗粒,并隔离mRNA和调节蛋白。
(5)These findings describe a cytoplasmic osmosensing mechanism that could allow plants to rapidly adjust cellular homeostasis under osmotic stress.这些发现描述了一种细胞质渗透感应机制,这种机制可以使植物在渗透胁迫下快速调节细胞稳态。
Structured Abstract
INTRODUCTION
(1)Living with water, cells are frequently challenged by osmotic perturbations微扰. The imbalance between the osmotic pressures across the semipermeable plasma半透膜血浆 membrane forces water to move in or out of a cell (through a process known as osmosis), remolds重塑 its shape, and can have substantial effects on various cellular activities. 生活在水中,细胞经常受到渗透扰动的挑战。半透性质膜两侧渗透压之间的不平衡迫使水进出细胞(通过一种称为渗透的过程),重塑其形状,并对各种细胞活动产生实质性影响。
(2)To preserve appropriate water and to maintain a suitable size, cells must sense and adapt to osmotic changes within their surrounding environments. 为了保持适当的水分和大小,细胞必须感知和适应周围环境的渗透变化。
(3)This is particularly true for most plant cells because they are directly exposed to the fluctuations波动 of environmental osmolarity. 对于大多数植物细胞来说尤其如此,因为它们直接暴露于环境渗透压的波动。
(4)For example, the root cells of land plants have to face osmotic stresses generated from dramatic changes of soil moisture, temperature, and salinity, which are major threats to agricultural production. 例如,陆地植物的根细胞必须面对由土壤水分、温度和盐度的剧烈变化产生的渗透胁迫,这是农业生产的主要威胁。
(5) Over the past decades, great efforts have been made to understand the adaptations of plants to such osmotic stresses, although how environmental osmotic changes are sensed by plant cells is far from fully understood.在过去的几十年里,人们已经做了大量的努力来了解植物对这种渗透胁迫的适应,尽管植物细胞是如何感知环境渗透变化的还远未完全了解。
(6)RATIONALE
(7)As a direct consequence of osmosis渗透, cell volume changes immediately upon nonisosmotic exposure. 作为渗透作用的直接结果,细胞体积在非等渗条件下会立即发生变化。
(8)Volumetric changes then trigger a series of physicochemical物理化学 alterations that could be perceived by the cells as osmotic signals. 体积的变化会引发一系列的物理化学变化,这些变化可以被细胞感知为渗透信号。
(9)A well-accepted example is that volumetrically turned membrane tension紧张 could be recognized by transmembrane mechanosensitive ion channels跨膜机械敏感离子通道, which represents a canonical 典型的osmosensing strategy. 一个广为接受的例子是,体积调节的膜张力可以被跨膜机械敏感离子通道识别,这代表了一种典型的渗透感应策略。
(10)In addition to membrane tension restricted to the plasma membrane, cell volume changes also have additional effects spreading all over a cell, such as changing intracellular molecular crowding states. 除了限制在质膜上的膜张力之外,细胞体积的变化还具有遍布整个细胞的额外效应,例如改变细胞内的分子拥挤状态。
(11)Thus, it raises an open question as to whether cellular osmosensing could be achieved elsewhere and through distinct mechanisms.因此,它提出了一个公开的问题,即细胞渗透感应是否可以在其他地方通过不同的机制实现。
(12)RESULTS
(13)First, we found that Arabidopsis Decapping 5 (DCP5) protein rapidly and reversibly可逆assembles to cytoplasmic condensates 细胞质凝聚物in response to hyperosmotic高渗的 stresses. 首先,我们发现拟南芥去盖蛋白5 (DCP5)在高渗胁迫下快速可逆地组装成细胞质浓缩物。
(14)This behavior is independent of the type of stress-triggering osmolytes and is not regulated by canonical osmosignaling pathways. .这种行为不依赖于压力触发的渗透调节物质的类型,也不受规范的渗透信号通路的调节。
(15)By tracing cell volume during hyperosmotic treatment, we uncovered a causal relationship between osmotic cell volume changes and DCP5 condensation. 通过追踪高渗治疗期间的细胞体积,我们揭示了渗透细胞体积变化和DCP5浓缩之间的因果关系。
(16)Then, we found that DCP5 undergoes phase separation in response to molecular crowding both in vivo and in vitro and confirmed that this phase behavior of DCP5 depends on its intrinsically disordered region (IDR). 然后,我们发现DCP5在体内和体外都经历了响应分子拥挤的相分离,并证实了DCP5的这种相行为取决于其内在无序区(IDR)。
(17)Moreover, we found that DCP5 IDR harbors an intramolecular crowding sensor 分子内拥挤传感器(ICS) that senses hyperosmotically intensified加强 molecular crowding through conformational构象的 change and triggers phase separation through hydrophobic interactions疏水相互作用. 此外,我们发现DCP5 IDR拥有一个分子内拥挤传感器(ICS ),它通过构象变化感知高渗强化的分子拥挤,并通过疏水相互作用触发相分离。
(18)This ICS is land plant–specific and does not exist in yeast, algae, or animal DCP5 homologs. On the other aspect, by uncovering mRNA and translation regulators as its components, DCP5 condensate凝结物 was identified as a distinct type of plant stress granule, which we designated as DCP5-enriched osmotic stress granule (DOSG). 这种ICS是陆地植物特有的,不存在于酵母、藻类或动物DCP5同系物中。另一方面,通过发现mRNA和翻译调节因子作为其成分,DCP5冷凝物被鉴定为一种独特类型的植物胁迫颗粒,我们将其命名为富含DCP5的渗透胁迫颗粒(DOSG)。
(19)We also confirmed that mRNA is recruited to DOSGs through the interaction between DCP5 and polyadenylate-binding proteins (PABs). 聚腺苷酸结合蛋白 我们还证实,mRNA通过DCP5和聚腺苷酸结合蛋白(PAB)之间的相互作用募集到DOSGs。
(20)Throughsequestration of plenty of mRNA and regulatory proteins at DOSGs, and a translation repression function of itself, DCP5 reprograms both the translatome and transcriptome from growth promotion to stress adaptation. 通过在DOSGs上隔离大量的mRNA和调节蛋白,以及自身的翻译抑制功能,DCP5将翻译组和转录组从生长促进重编程为应激适应。
(21)Accordingly因此, a DCP5-deficient mutant was hypersensitive to hyperosmotic stress and desiccation脱水, which could be completely rescued by full-length DCP5 but only partially recovered by ICS-deleted DCP5 that is disabled in phase separation and DOSG assembly因此,DCP5缺陷型突变体对高渗胁迫和脱水高度敏感,它可以被全长DCP5完全拯救,但只能被在相分离和DOSG装配中失活的ICS缺失型DCP5部分恢复。
(22)CONCLUSION
(23)Our findings uncovered a cytoplasmic osmosensing and stress adaptation mechanism mediated by molecular crowding–triggered phase separation of DCP5 and DOSG assembly, identified DCP5 as a multifunctional osmosensor that participates in both osmosensing and osmotic stress adaptation, and suggested a stress sensory function for hyperosmotically induced stress granules. 我们的发现揭示了一种由分子拥挤触发的DCP5和DOSG装配相分离介导的细胞质渗透感应和压力适应机制,鉴定了DCP5是一种多功能渗透传感器,参与渗透感应和渗透压力适应,并提示了高渗诱导的压力颗粒的压力感觉功能。
Words:
osmolarity 渗透压
hyperosmolarity 高渗性
sequesters 隔离
molecular crowding
granules 微粒
remolds 重塑
cytoplasmic condensates 细胞质凝聚物